The Brushy Mountains Brood
Marlatt’s map of Brood VI (Marlatt 1923) has always been problematic since it suggests that populations of this brood are present across the general range of periodical cicadas. Marlatt’s map shows populations scattered over eastern North America, including extensive areas of emergence in the Philadelphia-New York corridor and in the Midwest. Some Marlatt records, such as those in Wisconsin, are supported by museum specimens (University of Wisconsin-Madison), though as with many older museum collections, specific information about date and locality of collection is lacking. Simon’s (Simon 1988) revised map of Brood VI shows all but the “core” populations in the mountains of North Carolina, South Carolina, and Georgia as doubtful. This brood includes the species Magicicada septendecim, and Magicicada septendecula; M. cassini is absent.
In the map below, cicada symbols are verified presence records and red symbols are verified absence records in our database as of March 2025. Click on points for details. Gold symbols are from Simon (1988); smaller symbols are records with a lower degree of certainty and black crosses represent records that are considered spurious. Blue symbols are from Marlatt (1923); smaller symbols are records with a lower degree of certainty and question marks represent records that are considered spurious. Symbols are in layered in the order Database, Simon, Marlatt, and symbols in the upper layers may obscure symbols in lower layers. Some absence records in the database are not shown for clarity. Note that the dates shown do not represent dates of adult emergence; rather, they represent dates on which choruses were active. Thus, in any given area, adult emergence may have occurred a week or more earlier than the dates shown on this map. This map may not be reproduced without written permission.
Marshall (2001) noted that many Brood VI records may be the products of biases in earlier mapping efforts; some biases have to do with the ways that records were compiled over generations, while others have to do with varying degrees of effort to search for emergences. A key source of bias is the practice of assigning records to broods on the basis of expectation and timing, a problem compounded by a lack of emergence density data associated with such records. Periodical cicadas have a propensity to emerge off-schedule or “straggle” in increments of four years (Lloyd and Dybas 1966, Dybas 1969, Lloyd and White 1976, Simon and Lloyd 1982, Maier 1985, Kritsky 1988, Kritsky and Young 1992, Marshall et al. 2011). When broods in close physical proximity (such as II, VI, X, and XIV; Cooley et al. 2009, Cooley et al. 2011, Cooley et al. 2015) are offset by increments of 4 years, the brood membership of low-density populations or populations along the periphery of any one brood can be uncertain. Emerging cicadas could belong to the brood emerging in a particular year, or they could be a “shadow brood” –stragglers from the temporally and spatially related brood; moreover, such populations could be self-sustaining, or they could persist only because they are continually replenished by more stragglers (Lloyd and White 1976, Marshall 2001, Cooley et al. 2011). Shadow brooding seems to be the best explanation for records of “Brood XV” which, if it existed, would emerge four years before Brood II (Lloyd and White 1976, Simon and Lloyd 1982, Cooley et al. 2011). Maier’s (Maier 1985) reports of cicadas in Connecticut emerging on the same schedule as Brood VI may have a related explanation, as they emerged four years after Brood II in the same areas. Because there are so many possibilities for shadow brooding involving Brood VI, many of the records in Marlatt (Marlatt 1923) attributed to Brood VI may actually belong to other broods –they could just be “shadows” of them (Marshall, 2001). Other records, such as those in Wisconsin, are not easily explainable as belonging to other broods, because they are nowhere near likely sources of stragglers (e.g., II, X, or XIV). Thus, the temporal and spatial context of Brood VI makes it especially challenging to map. In addition, interactions between neighboring broods offset by increments of four years may create the conditions in which one brood replaces another over time (Lloyd and White 1976, Kritsky 1988, Kritsky and Young 1992).
Our project has constructed detailed maps of Brood VI in two generations (Cooley et al. 2021). We mapped the main body of Brood VI in 2000, marking paper maps and later transcribing them:
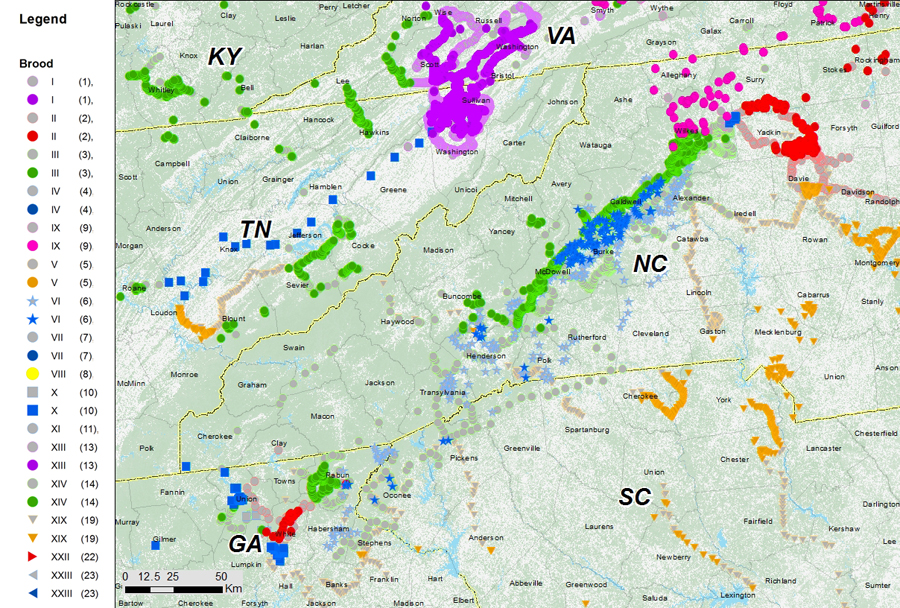
In 2017, we mapped the brood using GPS dataloggers. Overlaying the records from the two generations suggests that in some places, Brood VI expanded its range slightly between 2000 and 2017, although the measured changes are only on the order of 1-2km (Cooley et al. 2021). We found no evidence of Brood VI in Wisconsin in either year.